2020-09-08 理化学研究所,岡山大学
理化学研究所(理研)環境資源科学研究センターバイオ生産情報研究チームの香西雄介研究員、持田恵一チームリーダーと岡山大学大学院環境生命科学研究科(農)の能年義輝准教授らの共同研究チームは、農作物の重要病害である紋枯病[1](もんがれびょう)から植物が身を守る仕組みを明らかにしました。
本研究成果は、紋枯病に対する新たな防除策を開発するための分子基盤となるものと期待できます。
紋枯病は、イネやトウモロコシなどの穀物に大きな被害をもたらす重要病害ですが、植物が菌の感染に抵抗する機構はよく分かっていませんでした。
今回、共同研究チームは、モデル植物であるミナトカモジグサ[2]において、紋枯病菌に対して抵抗性を示す系統と罹病性を示す系統を用い、それぞれに菌を感染させ、その後の遺伝子発現変動を経時的かつ網羅的に比較解析しました。その結果、抵抗性を示す系統は菌に対する免疫応答を素早く活性化させていることが分かりました。また、機械学習[3]を用いた解析によりこの免疫応答の転写調節に関わる制御因子を探索し、BdWRKY38転写因子[4]の同定に成功しました。さらに、BdWRKY38遺伝子を発現させると、罹病性系統は紋枯病抵抗性を獲得することを確認しました。
本研究は、科学雑誌『The Plant Journal』の掲載に先立ち、オンライン版(9月5日付)に掲載されました。
背景
紋枯病(もんがれびょう)は、イネやトウモロコシなど穀物の重要病害であり、土壌に生息する植物病原糸状菌Rhizoctonia solaniによって引き起こされます。紋枯病は、栽培イネではいもち病に次ぐ二大病害の一つであり、最大で50%もの収量減をもたらします。現在、紋枯病に対して強い抵抗性を示す栽培イネの品種・系統は見いだされておらず、農業現場では栽培管理や殺菌剤を用いた防除に頼っています。殺菌剤に耐性を持つ紋枯病菌が報告されたことや、地球環境変動に伴う今後の被害拡大が懸念されていることから、紋枯病に対する新たな防除策の開発が求められています。
共同研究チームは、これまでにイネ科のモデル草本植物[5]であるイネとミナトカモジグサを用いて、植物と紋枯病菌の相互作用について研究を進めてきました。2017年には、植物がホルモンの一つであるサリチル酸[6]を介した免疫機構により紋枯病抵抗性を発揮する能力を持つこと、またイネとは異なり、ミナトカモジグサには紋枯病に強い自然系統が存在することを見いだしました注1)。
ミナトカモジグサの紋枯病抵抗性系統では、菌の感染によってBdWRKY38とBdWRKY44という二つのWRKY転写因子[7]をコードするサリチル酸応答性遺伝子が誘導されます。これらのことから、元来紋枯病に対する抵抗性を備えている植物は、サリチル酸を介した免疫を駆使して菌の感染を防いでいると予想されていましたが、その具体的な仕組みはよく分かっていませんでした。
そこで、共同研究チームは、抵抗性の強弱が異なるミナトカモジグサ系統の紋枯病菌に対する応答を詳しく比較することにより、植物が菌の感染に抵抗する機構を調べました。
注1)2017年10月20日岡山大学プレスリリース「イネやバイオマス作物を脅かす紋枯病から身を守るための植物免疫機構を解明」
研究手法と成果
共同研究チームはまず、紋枯病に強いミナトカモジグサ系統のBd3-1を用いて、サリチル酸、およびサリチル酸応答性のWRKY転写因子であるBdWRKY38とBdWRKY44が、その抵抗性に担う役割について調べました。細菌由来のサリチル酸分解酵素遺伝子(NahG)を導入して内生のサリチル酸を減少させた植物と、BdWRKY38遺伝子の発現をRNA干渉法(RNAi法)[8]により抑制した植物では、野生型のBd3-1と比べて紋枯病抵抗性が著しく低下しました(図1)。BdWRKY44遺伝子の発現抑制植物でも抵抗性の低下は認められましたが、その影響は小さいものでした。つまり、Bd3-1が自然に備えている紋枯病抵抗性には、サリチル酸とその下流のBdWRKY38転写因子が重要な役割を果たしていることが分かりました。

図1 サリチル酸応答性のWRKY遺伝子の発現を抑制した植物の紋枯病抵抗性
ミナトカモジグサBd3-1は紋枯病菌に対して抵抗性を示すが、BdWRKY38遺伝子の発現を抑制、またはサリチル酸分解酵素遺伝子(NahG)を強発現させると、その抵抗性は著しく低下した。一方、BdWRKY44遺伝子の発現抑制では、抵抗性はあまり低下しなかった。
次に、抵抗性の強弱が異なるミナトカモジグサ系統を用いて、紋枯病菌の感染過程を観察しました。罹病性系統のBd21と比べると、抵抗性系統のBd3-1とTek-3では、菌を接種した葉における菌糸量の増大や、宿主細胞へ侵入するための感染器官の形成がほとんど認められず、菌の感染が早期に食い止められていることが分かりました。
そこで、これらの系統の紋枯病菌に対する防御応答を比較するため、菌を感染させた後の時系列比較トランスクリプトーム解析[9]を行いました。網羅的な遺伝子発現パターンと遺伝子オントロジーエンリッチメント解析[10]の結果、抵抗性のBd3-1とTek-3では、防御応答遺伝子群が速やかに発現誘導されるのに対し、罹病性のBd21では遅れて誘導されることが分かりました(図2)。また、WRKY転写因子をコードする遺伝子群およびWRKY転写因子が結合する配列(W-box)をプロモーター領域[11]に持つ遺伝子群の発現パターンの解析から、ミナトカモジグサは紋枯病菌の感染に対して、WRKY転写因子により制御される防御応答によって対抗しており、抵抗性系統ではその活性化が早く起こることが分かりました。
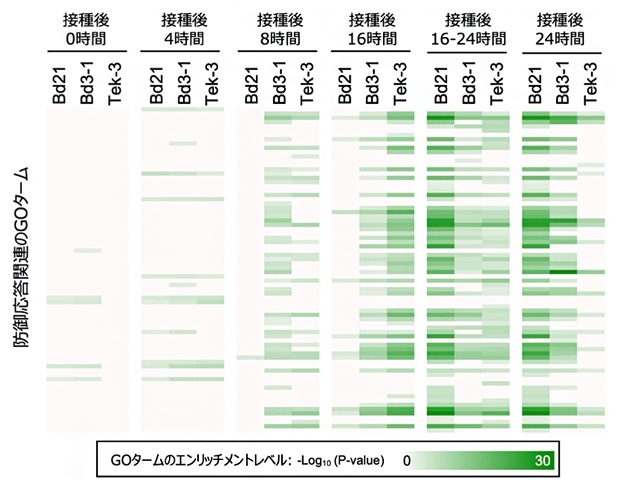
図2 紋枯病菌を感染させた植物における防御応答遺伝子群の発現パターン
防御応答遺伝子群の発現パターンを遺伝子オントロジー(GO)エンリッチメント解析により間接的に調べた。抵抗性のBd3-1とTek-3では防御応答遺伝子群が菌の接種後8~16時間で発現誘導されるのに対し、罹病性のBd21では遅れて接種後16~24時間で誘導されていた。
次に、機械学習を用いた遺伝子制御ネットワーク[12]解析により、ミナトカモジグサの紋枯病菌に対する防御応答をさらに解析しました。抵抗性のBd3-1とTek-3と罹病性のBd21におけるネットワーク構造を比較したところ、BdWRKY44は全ての系統のネットワークでハブを構成していましたが、BdWRKY38は抵抗性系統に特異的なハブであることが分かりました。つまり、BdWRKY38を駆動できることが、紋枯病抵抗性の鍵であることを示しています。これを確かめるために、罹病性系統のBd21においてBdWRKY38を強く発現する系統を作出したところ、野生型のBd21と比べて紋枯病抵抗性が高まっていることが分かりました。
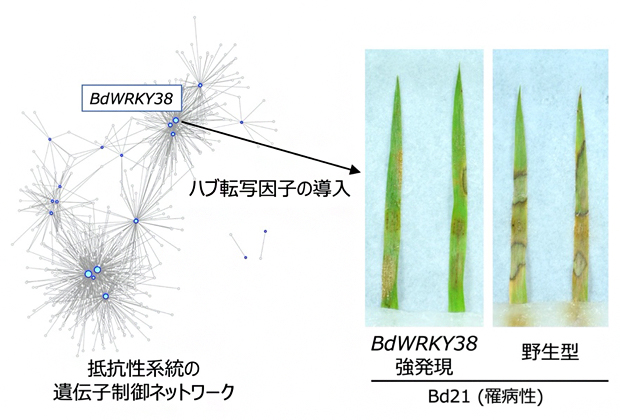
図3 BdWRKY38遺伝子を強発現する罹病性植物の紋枯病抵抗性
ミナトカモジグサBd21は紋枯病菌に対して罹病性を示すが、抵抗性系統の遺伝子制御ネットワークでハブを構成するBdWRKY38遺伝子を強発現させると、病徴が軽減された。
今後の期待
本研究により、ミナトカモジグサが自然に備えている紋枯病抵抗性機構の一端が明らかになりました。防御関連遺伝子群の素早い発現誘導、菌増殖の早期の抑制、抵抗性の発現へのサリチル酸の必要性などの特徴は、一般的に植物が非親和性の病原体に対して示す防御応答とよく似ていました。すなわち、ミナトカモジグサは、イネが持たない紋枯病菌を感知するためのセンサーとして働く抵抗性遺伝子を保持していることを示唆しています。今後はその遺伝子資源を特定し、病害防除へ応用することを目指しています。
なお本研究は、植物病害被害の軽減を通じ、国際連合が掲げる「持続可能な開発目標(SDGs)[13]」のうち「2.飢餓をゼロに」及び「15.陸の豊かさも守ろう」への貢献が期待される成果といえます。
補足説明
1.紋枯病
植物病原性の糸状菌Rhizoctonia solaniが原因となる栽培イネの重要病害で、トウモロコシやソルガムといった他の穀物にも発生する。栽培イネにおける第一次伝染源は菌核であり、水田の灌水などによりイネの葉鞘に付着し、感染が始まる。感染部位やその周辺には灰色で楕円形の紋型病斑が形成され、植物体の倒伏、枯死、米粒の品質低下などをもたらす。
2.ミナトカモジグサ
イネ科イチゴツナギ亜科に属する小型の植物。実験室で扱いやすい特徴を備えており、ゲノム情報も整備されているため、モデル草本植物として利用されている。学名はBrachypodium distachyon(ブラキポディウム・ディスタキオン)。
3.機械学習
コンピューターを用いて、数値や画像などの膨大なデータから有用な知識やルールを見つけ出す技術の総称。
4.転写因子
DNA上に存在する遺伝子の転写調節領域に結合し、DNAからmRNAへの転写量やタイミングを調節するタンパク質。
5.草本植物
地上部にある茎の生存期間が短く、木化や肥大成長することがほとんどない植物。イネやコムギなど穀物として利用されるイネ科植物のほとんどが含まれる。
6.サリチル酸
植物ホルモンの一種であり、細菌、糸状菌、卵菌、ウイルスなどに対する植物の病害抵抗性において重要な役割を果たす。
7.WRKY転写因子
植物に特有な転写因子ファミリーの一つ。プロモーター領域にW-box配列を持つ遺伝子の発現を制御する。
8.RNA干渉法(RNAi法)
短い2本鎖RNAの導入によって、それと相補的な配列を持つmRNA特異的に分解され、遺伝子発現が抑制される現象。RNAiは、RNA interferenceの略。
9.トランスクリプトーム解析
ある状態の生物における全遺伝子の発現レベルを網羅的に解析する手法。ここでは、菌を感染させた後の時系列サンプルを扱い、罹病性と抵抗性植物の相違を比較していることから、時系列比較トランスクリプトーム解析としている。
10.遺伝子オントロジーエンリッチメント解析
遺伝子に付与される注釈のことを遺伝子オントロジー(Gene ontology, GO)といい、biological process(生物学的プロセス)、cellular component(細胞の構成要素)、molecular function(分子機能)に分類される。遺伝子オントロジーエンリッチメント解析は、ある遺伝子セットの中に、特定の遺伝子オントロジーを持つ遺伝子が過剰に含まれているか否かをエンリッチメント解析により調べる手法。
11.プロモーター領域
遺伝子の上流に存在する転写因子が結合するDNA配列を含む領域のことで、遺伝子の発現量や発現タイミングの制御を担う。
12.遺伝子制御ネットワーク
転写因子とその制御を受ける被制御遺伝子との関係性を描画したネットワーク。ここでは、遺伝子発現量データセットから決定木ベースの機械学習ツールを用いて、制御関係を推定している。ネットワーク上で多数の遺伝子と結合する遺伝子(ハブ遺伝子)は、生物学的に重要であると考えられている。
13.持続可能な開発目標(SDGs)
2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された2016年から2030年までの国際目標。持続可能な世界を実現するための17のゴール、169のターゲットから構成され、発展途上国のみならず,先進国自身が取り組むユニバーサル(普遍的)なものであり、日本としても積極的に取り組んでいる(外務省ホームページから一部改変して転載)。
共同研究チーム
理化学研究所
環境資源科学研究センター バイオ生産情報研究チーム
研究員 香西 雄介(こうざい ゆうすけ)
テクニカルスタッフ 清水 みなみ(しみず みなみ)
テクニカルスタッフ 井上 小槙(いのうえ こまき)
テクニカルスタッフ 上原(山口) 由紀子(うえはら(やまぐち) ゆきこ)
大学院生リサーチ・アソシエイト 高萩 航太郎(たかはぎ こうたろう)
パートタイマー 中山 梨沙(なかやま りさ)
チームリーダー 持田 恵一(もちだ けいいち)
岡山大学
資源植物科学研究所
技術専門職員 松浦 恭和(まつうら たかかず)
准教授 森 泉(もり いずみ)
教授 平山 隆志(ひらやま たかし)
大学院 環境生命科学研究科(農)
大学院生 ソブヒ・アブデルサラム(Sobhy S.H. Abdelsalam)
准教授 能年 義輝(のうとし よしてる)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費補助金研究活動スタート支援「ミナトカモジグサを用いた紋枯病抵抗性機構の解明とその原因遺伝子の同定(領域代表者:香西雄介)」、若手研究B「紋枯病に対する植物の抵抗性発現機構の分子基盤(領域代表者:香西雄介)」、岡山大学資源植物科学研究所共同研究課題「ミナトカモジグサ突然変異体リソースを用いた病害罹病性遺伝子の探索(領域代表者:香西雄介)」による支援を受けて行われました。
原論文情報
Yusuke Kouzai, Minami Shimizu, Komaki Inoue, Yukiko Uehara-Yamaguchi, Kotaro Takahagi, Risa Nakayama, Takakazu Matsuura, Izumi C. Mori, Takashi Hirayama, Sobhy S.H. Abdelsalam, Yoshiteru Noutoshi, Keiichi Mochida, “BdWRKY38 is required for the incompatible interaction of Brachypodium distachyon with the necrotrophic fungus Rhizoctonia solani”, The Plant Journal, 10.1111/tpj.14976![]()
発表者
理化学研究所
環境資源科学研究センター バイオ生産情報研究チーム
研究員 香西 雄介(こうざい ゆうすけ)
チームリーダー 持田 恵一(もちだ けいいち)
岡山大学 環境生命科学研究科(農)
准教授 能年 義輝(のうとし よしてる)
報道担当
理化学研究所 広報室 報道担当
岡山大学 総務・企画部広報課

