2024-07-09 明治大学
研究成果のポイント
- 種子発芽の時季を左右する「発芽の温度範囲」を決める仕組みに、細胞内情報伝達経路、“MKK3-MAPキナーゼカスケード”が働くことを発見しました。
- MKK3-MAPキナーゼカスケードは、種子の休眠状態と環境の温度の組み合せで活性化され、植物ホルモン作用の制御を介して発芽の温度範囲を広げ、休眠性の低下をもたらすことを明らかにしました。
- MKK3-MAPキナーゼカスケードは、作物種子の品質を左右する収穫前の「穂発芽」や、生産効率を低下させる高温や低温による発芽阻害を制御する技術開発のターゲットになると期待されます。
- MAPキナーゼは、昆虫(カイコガなど)では低温による胚休眠の解除に働くことが示されており、生物が温度を感知して休眠を制御する仕組みの解明につながることが期待されます。
研究の概要
種子が発芽する季節・タイミング(時季)は、種子自身が持つ休眠の状態注1と環境の温度の組合せによって決まります。種子成熟後の時間経過に伴う休眠性の低下は、発芽可能な温度範囲の拡大をもたらし、環境の温度がこの範囲に収まる時季に発芽します(図1)。種子は、本来と異なる時季に発芽してしまうと、個体の成長や種子生産がダメージを受けるため、生育に適したタイミングで発芽することが重要です。今回、明治大学農学部の川上直人教授、明治大学大学院農学研究科博士後期課程の大谷真彦(現:博士(農学)、明治大学研究・知財戦略機構 研究推進員)をはじめとする国際共同研究グループ(パリ-サクレー大学・フランス国立農業食料環境研究所のJean Colcombet博士、香川大学農学部の市村和也教授、国立遺伝学研究所の越水静助教ら)は、種子が発芽可能な温度範囲を決める仕組みを解明しました。この仕組みでは、酵母から植物、動物まで広く存在する細胞内情報伝達経路、MAPキナーゼカスケード注2が中心的な役割を持ち、発芽の適温でカスケードが活性化され、植物ホルモンの作用を調節して発芽を促し、発芽の温度範囲を決めていることを明らかにしました。このカスケードで働くタンパク質リン酸化酵素(キナーゼ)の一つ、MKK3は、コムギやオオムギでは小麦粉やモルト(麦芽)の品質を大きく左右する穂発芽関連遺伝子です。今回の研究は、種子が温度を感知して発芽の時季を制御する仕組みの解明につながるだけでなく、温暖化に対応し、安定した作物生産を可能とする、種子発芽制御技術の開発への応用が期待されます。
本研究成果は、米国科学アカデミーが発行する総合科学学術誌「Proceedings of the National Academy of Sciences of the United States of America(PNAS)」に掲載されました。
研究の背景
種子が発芽する時季の決定は、その後の個体の成長のみならず、種の存続をも左右します。たとえば、暑さに弱い植物が春に芽生えてしまうと、花を付ける前に枯れてしまったり、花は咲いても種子が実らなかったりします。種子は、たとえ発芽に適切な環境でも発芽を抑える「休眠性」を持ち、環境(土壌)の温度を感知することにより、生育に不適な季節の発芽を抑え、適切な季節の発芽を実現しています。たとえば、秋に発芽して春に種子を付ける「冬型一年生草本」の種子は、春から秋にかけて、休眠性の低下と共に発芽可能な温度の上限を上昇させ、土壌温度が上限を下回る秋に発芽します(図1)。この場合、休眠性の低下は、発芽可能な温度範囲の拡大と発芽至適温度の上昇によってもたらされます。もし、発芽可能な温度範囲が狭まると、発芽の時季がずれ、期間も限定されてしまうでしょう(図1:実線から点線、緑から紫の矢印へ)。これまでに、発芽の至適温度を制御する遺伝子は見出されていますが、発芽可能な温度範囲を調節する仕組みは知られていませんでした。

図1 冬型一年生草本種子の発芽の季節決定:種子の休眠と環境の温度
秋に発芽する冬型一年生草本の種子は春に成熟して地上に散布される。このとき、種子を様々な温度で吸水させても発芽しない。春から秋にかけて、種子の休眠性の低下と共に、発芽可能な温度の上限が上昇して発芽可能な温度範囲(緑の網掛け部分)が拡大し、環境の温度が上限を下回る秋に発芽する。
ところで、私たち人類にとって種子は保存が効く食料であり、安定した作物生産の出発点です。ところが、収穫前の降雨や多湿な環境によって親植物上で種子が発芽してしまう「穂発芽」は、種子の品質や収量を大きく低下させます(図2)。発芽した種子は乾燥耐性を持たないため、収穫した種子の発芽率は低下し、種子から加工した製品の品質は大きく低下します。近年、コムギとオオムギに穂発芽をもたらす原因遺伝子として、MAPキナーゼカスケード注2を構成するタンパク質リン酸化酵素をコードする、MKK3が同定されました。オオムギでは、MKK3活性を低下させる変異が種子の休眠性を高め、穂発芽耐性をもたらすことが示されています。しかし、MKK3が他の多くの植物でも休眠と発芽に作用するかは知られていませんでした。また、MAPキナーゼカスケードの全貌、つまりMKK3を活性化するMAPKKK、MKK3が活性化するMPKは同定されておらず、MAPキナーゼカスケードがどのような条件で活性化され、どのようにして休眠性を制御しているかも未解明でした (図2) 。
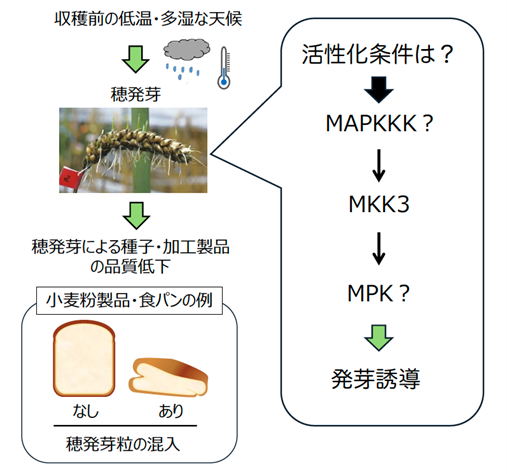
図2 穀類の穂発芽耐性に品種間差をもたらすMKK3遺伝子
収穫前の多湿な環境では、種子が母体についたまま発芽してしまう「穂発芽」が誘導されることがある。コムギでは、穂発芽粒を含んだまま製粉してしまうと、デンプン等が分解され、加工食品の品質が劣化してしまう。穀類の穂発芽耐性に品種間差をもたらす遺伝子の一つとしてMKK3が同定されている。
研究体制
【国際共同研究グループ】※所属は研究当時の内容
明治大学 農学部生命科学科 植物分子生理学研究室
教授 川上直人
農学研究科博士後期課程 大谷真彦
農学研究科博士前期課程 東城僚
農学研究科博士後期課程 鄭李鵬
農学研究科博士前期課程 星拓実
農学研究科博士前期課程 大森涼葉
農学研究科博士後期課程 橘夏希
農学研究科博士前期課程 佐野智広
パリ-サクレー大学・フランス国立農業食料環境研究所 博士 Jean Colcombet
パリ-サクレー大学大学院博士後期課程 Sarah Regnard
国立遺伝学研究所 助教 越水静
香川大学農学部 教授 市村和也
本研究の一部は、文部科学省私立大学戦略的研究基盤形成支援事業(S1411023)、明治大学新領域創生型研究(#229831002)の支援を受けて行われたものです。
研究成果
コムギやオオムギと進化的に離れたシロイヌナズナを材料として、MKK3遺伝子の機能が失われた突然変異、mkk3種子の発芽を調べたところ、正常な野生型種子よりも休眠性が強くなっていました。このため、MKK3は種を越えて種子の休眠を弱め、発芽を促す機能を持つことが分かりました。さらに、収穫直後と収穫後5ヶ月の休眠性が低下した種子(後熟種子)を用い、様々な温度で発芽を調べたところ、発芽可能な温度範囲が野生型よりも狭まっていることがわかりました。このことは、MKK3は種子が発芽できる温度範囲を拡大させる機能を持つことを示しています(図3)。
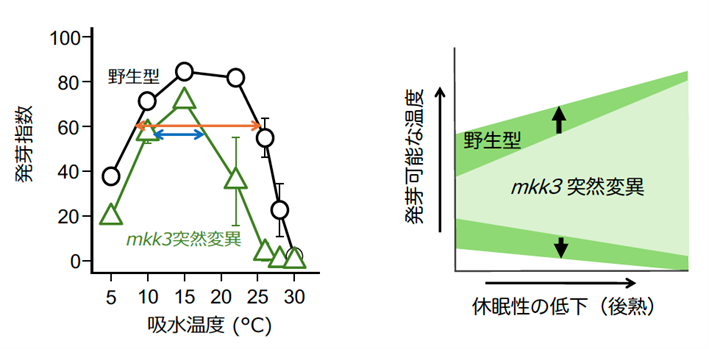
図3 MKK3は、発芽可能な温度範囲を拡大させる
左図の「発芽指数」は、発芽率と発芽速度の両者を加味し、発芽のしやすさを評価する指標。数値が高い方が発芽しやすいことを示す。mkk3突然変異体種子では、野生型種子よりも発芽可能な温度範囲(オレンジの矢印が野生型、青が突然変異体)が狭い。右図では、後熟に伴う発芽可能な温度範囲の拡大を模式的に表している。
シロイヌナズナは20のMPK遺伝子を持ちますが、MKK3によって活性化される可能性が報告されていたのは、MPK1, MPK2, MPK7, MPK14の4つでした。そこで、これらMPK遺伝子の機能喪失突然変異体種子の発芽を調べたところ、MPK7の機能喪失を含む多重変異体種子で明確な発芽率の低下が認められ、四重変異体の種子が最も発芽しにくくなりました。このため、いずれの遺伝子も休眠性を低下させ、高温条件で発芽を促す働きを持つこと、そして種子の休眠と発芽にはMPK7が主要に働くことが分かりました。シロイヌナズナにはMKK3の活性化に関わる可能性があるMAPKKK遺伝子が9つありますが、本研究ではこのうち2つの遺伝子の発現が発芽条件で誘導されることを見いだしました(図4a)。MAPKKK19とMAPKKK20遺伝子は、休眠性が低下した後熟種子を発芽の適温(26℃)で吸水させたときに、発現が強く誘導されました。一方、収穫直後の休眠種子が発芽しない26℃、そして後熟種子が発芽できない高温(34℃)では、両遺伝子とも発現が強く抑制されました(図4a)。さらに、この2つの遺伝子の機能が失われた二重変異体およびMAPKKK20遺伝子を過剰に発現する種子の発芽を調べ、MKK3遺伝子と同様、MAPKKK19とMAPKKK20遺伝子は休眠性を低下させ、発芽を促す働きを持つことを明らかにしました。
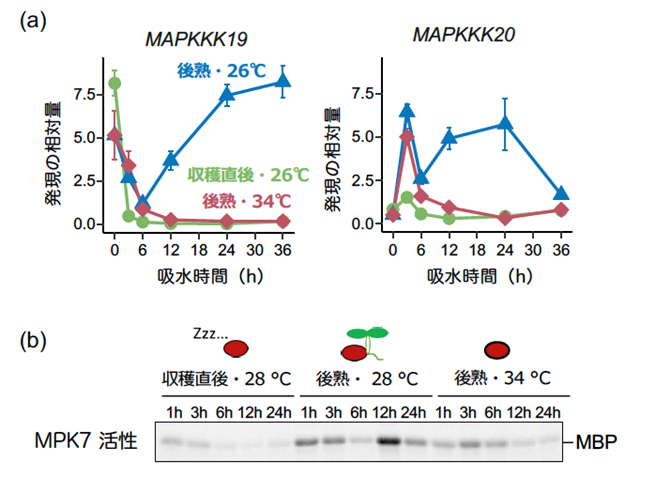
図4 種子の休眠性と温度がMKK3-MAPキナーゼカスケードの活性を制御する
(a) MAPKKK19とMAPKKK20遺伝子の発現を、発芽抑制条件(収穫直後の休眠種子を26℃、休眠性が低下した「後熟」種子を34℃で吸水)と、発芽誘導条件(後熟種子を26℃で吸水)で調べた。
(b) MPK7の活性化状態を、MAPキナーゼの基質としてよく利用されるMBPのリン酸化度合い(バンドの濃さ)から評価した。
さらに、MPK7のタンパク質リン酸化活性を生化学的に解析し、種子の休眠状態、環境の温度と、MKK3-MAPキナーゼカスケードの活性化の関連を調べました。MPK7はMAPKKK19/20遺伝子の発現に同調し、発芽が抑制される条件では活性化されず、後熟種子が発芽する温度条件でのみ活性化されました(図4b)。また、MKK3-MAPキナーゼカスケードは、発芽抑制に働く植物ホルモン、アブシシン酸の合成酵素遺伝子の発現を抑制し、分解酵素遺伝子の発現を誘導すること、発芽誘導に働く植物ホルモン、ジベレリン合成酵素遺伝子の発現を誘導する働きを持つことが分かりました(図5)。
今回の研究から、休眠性が低下した後熟種子が発芽の適温を感知すると、MAPKKK19/20遺伝子の発現を誘導することによりMKK3-MAPキナーゼカスケードを活性化し、植物ホルモンの作用制御を介して発芽を促すと考えられます(図5)。一方、収穫直後の種子は、後熟種子の発芽の適温(25℃前後)で吸水してもMAPKKK19/20遺伝子の発現が誘導されず、MKK3-MAPキナーゼカスケードの活性化が起こらないため、発芽できないと考えられます。
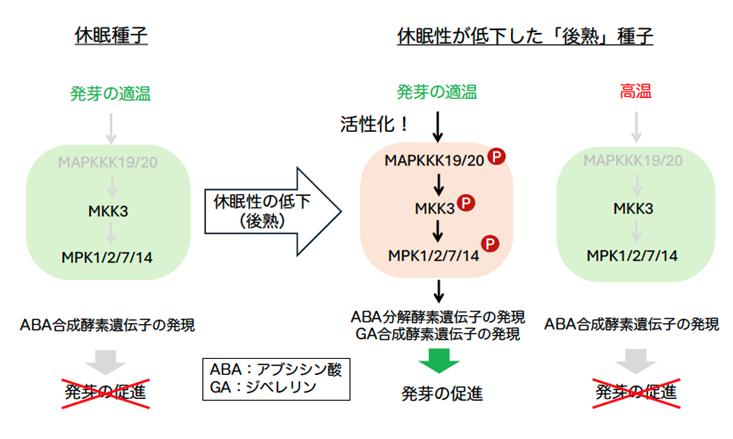
図5 MKK3-MAPキナーゼカスケードは種子の休眠状態と環境の温度情報を統合し、植物ホルモン代謝酵素遺伝子の発現制御を介して発芽を制御する
Pは、各キナーゼがリン酸化された活性化型であることを示す。
今後の期待
本研究では、周囲の温度を感知した種子が、MKK3-MAPキナーゼカスケードの活性を温度と休眠性の情報を統合して調節し、発芽を制御することを明らかにしました。今後は、種子が自身の休眠の状態と環境の温度を感知して、カスケード活性の制御に至るまでの仕組みを明らかにしたいと考えています。また、MKK3-MAPキナーゼカスケードから植物ホルモンの作用制御に至るまでの分子機構も解明したいと考えています。MAPキナーゼカスケードは、昆虫(カイコガなど)の卵(胚)の低温による休眠解除に働くことが示されています。今後、植物の種子はもちろん、生物が温度を感知して休眠を制御する仕組みの解明につながることを期待しています。
MKK3は穀類の穂発芽耐性に品種間差をもたらす遺伝子として発見されましたが、本研究はMKK3-MAPキナーゼカスケードが穀類以外の植物種で働いていることを示しています。穂発芽は穀類のみならず、野菜や果物などの種子でも見られ、播種後の発芽率にも大きく影響します。また、高温や低温による発芽阻害は生産効率を大きく低下させています。MKK3だけでなく、MKK3-MAPキナーゼカスケードを構成する遺伝子の変異を利用することにより、発芽形質の改善が期待されます。
論文情報
論文タイトル
The MKK3 MAPK cascade integrates temperature and after-ripening signals to modulate seed germination
著者
Masahiko Otani , Ryo Tojo , Sarah Regnard , Lipeng Zheng , Takumi Hoshi , Suzuha Ohmori , Natsuki Tachibana , Tomohiro Sano , Shizuka Koshimizu , Kazuya Ichimura , Jean Colcombet , Naoto Kawakami
掲載雑誌
Proceedings of the National Academy of Sciences of the United States of America (PNAS) 121巻28号 e2404887121
DOI
10.1073/pnas.2404887121
電子版公開日
2024年 7月5日(金)
URL
https://doi.org/10.1073/pnas.2404887121
用語説明
注1 種子の休眠と後熟
種子の休眠とは、たとえ発芽に適した光や温度、水分などの環境条件でも、生きた種子が発芽しない状態である。休眠は種子の成熟過程で獲得され、成熟後の時間経過や季節の変化を経て低下する。この過程を後熟と呼ぶ。後熟過程では発芽可能な温度範囲だけでなく、発芽可能な水分や光強度など範囲も拡大するが、その範囲を超えた条件では発芽せず、再び休眠を獲得する。
注2 MAP(分裂促進因子活性化タンパク質:Mitogen-Activated Protein) キナーゼカスケード
酵母から動物・植物まで進化的に広く保存された細胞内情報伝達経路の1つ。タンパク質リン酸化酵素(キナーゼ)であるMAPKKK、MKK、MPKから構成され、MAPKKKがMKK、MKKがMPKをそれぞれリン酸化により活性化し、細胞外の環境に対する応答に働くことが知られている。本研究の材料であるシロイヌナズナには80のMAPKKK、10のMKK、20のMPKが同定されており、植物の成長段階や多様な環境情報に対応したMAPKカスケードが働くと考えられている。
-
お問い合わせ先
研究に関するお問い合わせ
明治大学農学部生命科学科 教授 川上直人
香川大学農学部 教授 市村和也取材に関するお問い合わせ
明治大学 経営企画部 広報課
香川大学 企画総務部広報課
国立遺伝学研究所 広報室

")